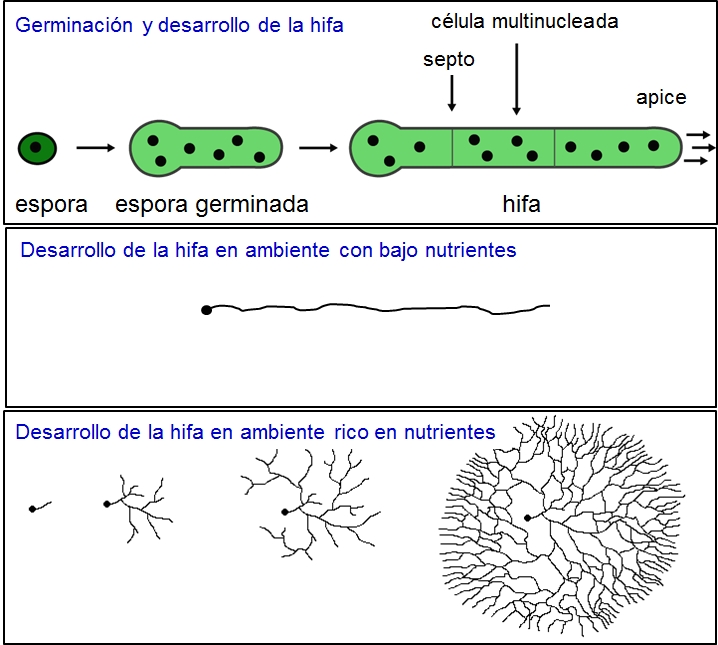
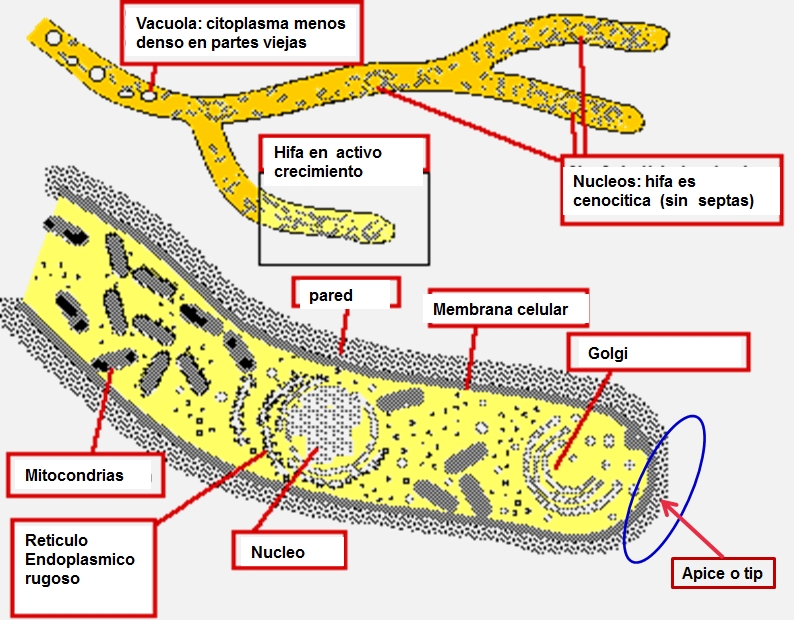
(Spoiler: Si)
Bien, veamos por qué

Personalmente me gusta consumir hongos, de diversas formas, así como los diversos hongos comercialmente disponibles. Pero este es un hábito no muy arraigado en la población, al menos latina. Las tendencias de consumo por filosofías naturistas, vegetarianas y veganas parecen estar cambiando e incrementando esta tendencia.
Para comenzar, los hongos tienen sus características organolépticas propias. Por ejemplo, el aroma, el olor a hongo es típico (algunos le dicen humedad) pero el sabor y la textura son también propios de ellos. Los hongos tienen un sabor particular, conocido como umami que significa delicioso en japonés. Aunque hablamos de características que son algo subjetivas, en general los hongos son sabrosos y versátiles para poder utilizarlos en diversas preparaciones culinarias.
Recolección: primera forma de obtener hongos para consumo

El consumo de hongos ha estado asociado a comunidades humanas desde muy antiguo, siendo la recolección la primera forma de obtención de ellos. En Europa principalmente, hay una larga tradición de recorrer bosques y campiñas para colectar hongos, tradición que se extendió a norte américa. Estudios arqueológicos han mostrado que este consumo es increíblemente antiguo y puede haber estado asociado a la evolución de las sociedades humanas. En efecto, en el sur de Chile, en la región de los lagos, se descubrió un antiguo asentamiento humano, en la localidad de Monte Verde, a 28 km de la ciudad de Puerto Montt (ver aquí detalles generales). Ahí se encontraron los vestigios de lo que podrían corresponder al asentamiento humano más antiguo del que se tiene registro en América (Aquí paper). En este yacimiento se hallaron objetos a los que se les estima una antigüedad de 14 mil años, entre ellos hongos que habría sido consumido por la comunidad de aborígenes. Ahora, parece ser que esta costumbre no trascendió mucho a las generaciones futuras, pues en Chile el consumo no es muy masivo, aunque en el último tiempo se puede apreciar un aumento de esta costumbre.
Producción comercial: actual forma para disponer variados hongos
Actualmente los hongos pueden cultivarse, bajo condiciones de temperatura, humedad y ciclos lumínicos, más un sustrato adecuado, lo que sustenta la producción comercial de diversos tipos de hongos. Hay varios países que tienen tradición de cultivo debido particularmente a su tradición de consumo. Sin embargo, es China el principal productor en cuanto a la cantidad producida. Le siguen países europeos, liderado por Italia, mientras que en américa están USA y Canadá. (Fuente).
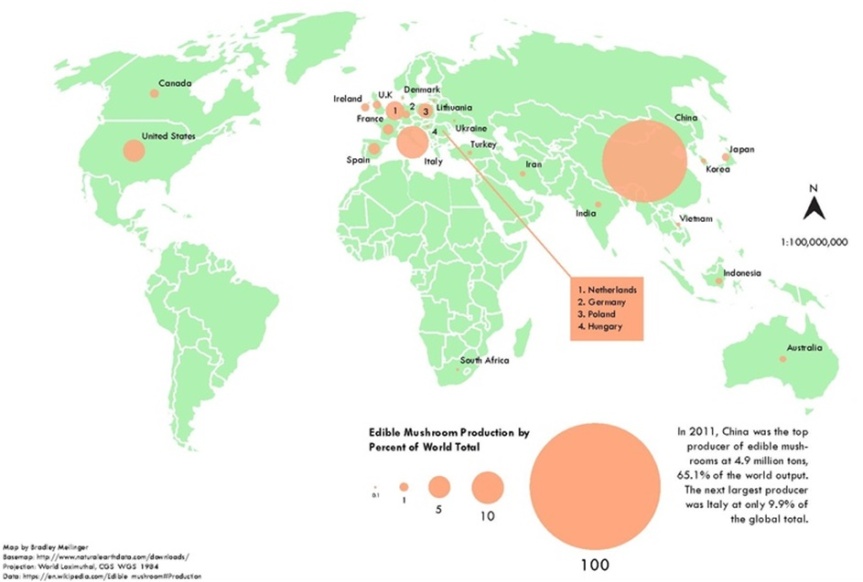
Producción mundial de hongos comestibles a nivel industrial
Dado al actual conocimiento, hay muchos hongos disponibles en el mercado. Entre los más comunes podemos mencionar al Champiñón de Paris, Portobello, Shiitake y el hongo ostra.

ASPECTOS NUTRICIONALES
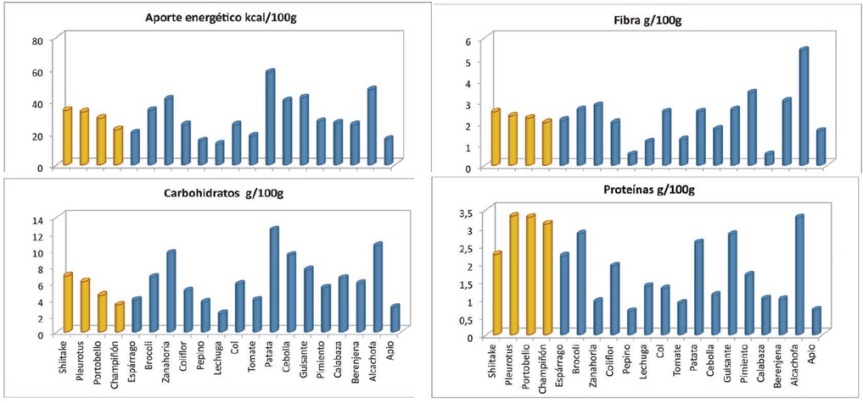
Los hongos son alimentos con unas propiedades nutricionales muy particulares que los hace muy apreciados entre los conocedores. Destaca el bajo aporte calórico que tienen debido a su gran contenido en agua (80%-90%) y a que sólo tienen entre 20 y 30 Kcal por cada 100 gramos dependiendo de la especie. Son una buena fuente de proteínas, con una composición en aminoácidos más parecida a la proteína animal que a la vegetal, siendo por lo tanto el complemento ideal para dietas vegetarianas y veganas. Su alto contenido en fibra y bajo aporte graso son características deseables para una alimentación saludable. En cuanto a los micro elementos, los hongos son una fuente importante de vitaminas del grupo B y de precursores de vitamina D como lo es el ergosterol. Contienen también minerales esenciales para el correcto funcionamiento de nuestro organismo, principalmente selenio, fósforo y potasio (ref) .
Aporte energético.
Los hongos proporcionan aproximadamente entre 26-35 kcal/100 g en función de la especie. El champiñón es uno de los que menos calorías aportan (26 kcal/100g) y el shiitake, aun siendo el hongo que mayor contenido energético presenta, sólo aporta 35 kcal.
Los hongos comestibles tienen un porcentaje muy alto de humedad (81,8-94,8%). El contenido en materia seca de champiñones es, por tanto, muy bajo, alrededor de 10%, y principalmente se compone de carbohidratos, proteínas, fibra y minerales.
Los hongos son alimentos con alto poder saciante y con baja densidad energética, que es la relación entre las calorías y el volumen de un alimento. El alto grado de humedad que tienen se relaciona con la sensación de saciedad que confieren y como además aportan muy pocas calorías, son un producto muy útil para dietas hipocalóricas.
Proteínas
El contenido proteico de los hongos oscila entre 15 y 35% de peso seco, dependiendo de las especies y el método de cultivo. Este contenido es bastante bueno y alto si compara con los contenidos de otros vegetales, tal como el 7 % en arroz, 9 % en maíz, 13 % en trigo y 38 % en soya. Por otro lado, la digestibilidad de esta proteína es en general bastante buena, por ejemplo, para el hongo ostra y shiitake es 73,4% y 76,3%, respectivamente.
A diferencia de las hortalizas, los hongos contienen todos los aminoácidos esenciales, que son aquellos que el cuerpo humano no puede generar por si solo y tiene que ingerirlos con la dieta. El género Pleurotus es el que mayor calidad proteica tiene ya que presenta una buena distribución de aminoácidos esenciales y no esenciales.
Carbohidratos.
Estos varían entre un 35 al 75% del peso seco de los hongos, según la especie. El shiitake (17,62 g/100g) es la que presenta mayor cantidad de carbohidratos totales comparada con el champiñón de parís (5,98 g/100g), con el hongo ostra (9,30 g/100g) y con el hongo del cardo (8,95 g/100g).
El contenido de azucares libres (mono y disacáridos) es bajo y puede ascender hasta el 11%. El manitol, representa uno de los principales azucares libres de los hongos. La glucosa es muy baja. De esta manera, no es un problema para diabéticas ya que no tiene impacto en el índice glicémico.
El mayor porcentaje de carbohidratos corresponde a polisacáridos complejos que otorgan un alto contenido de fibra soluble e insoluble. Estos tienen otras propiedades que revisare mas adelante en esta nota.
Fibra.
Los hongos son una buena fuente de fibra dietética, conteniendo más cantidad de fibra insoluble que de soluble. Los hongos que mayor porcentaje de fibra total presentan son el hongo del chopo (Agrocybe aegerita), el champiñón de París y el hongo ostra. Los polisacáridos que se encuentran en mayor proporción en la fibra del champiñón son los beta-glucanos (4-13% de la fibra dietética total).
Un estudio del Reino Unido mostró que 100 g de champiñones frescos aportarían entre 5% y 25% de la ingesta recomendada diaria de fibra (18 g fibra/día en Reino Unido).
Grasas.
El contenido de grasas es bastante bajo, valor que se encuentra alrededor del 5% ps. Dominan ácidos grasos insaturados, con un alto contenido en ácido linoleico. Los hongos no contienen colesterol, sino que a su vez poseen un alto contenido de ergosterol, el cual es precursor de la vitamina D.
El Shiitake tiene los mayores niveles de poliinsaturados y menor cantidad de saturados que el resto de los hongos estudiados. Las especies del género Pleurotus destacan por ser las que más ácidos grasos monoinsaturados presentan.
Minerales
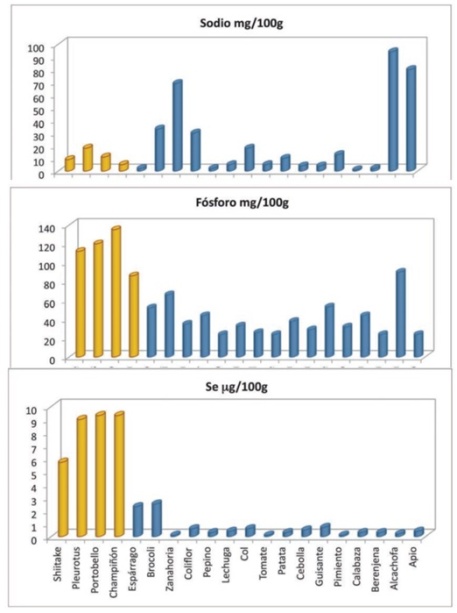
El contenido de minerales en los hongos varía entre 6 y 11% según la especie, por ejemplo, el hongo ostra tiene 6,90%, el del cardo 8,60%, el Shiitake 5,85% y la melena de león 9,35%.
Los macroelementos que más abundan en los hongos cultivados son calcio, fósforo, potasio y magnesio y de los microelementos destacan cobre, selenio, hierro y cinc.
En general, los hongos aportan muy bajo contenido de sodio por lo que son el alimento ideal para una dieta baja en sal.
Selenio. El contenido en selenio en los hongos varía de 0,46 a 5,63 ppm en función de la especie. Esta cantidad representa el 15% de la ingesta recomendada diaria en EEUU. Los hongos son mucho más ricos en selenio que las hortalizas y las frutas. Entre las funciones del selenio en nuestro organismo destaca su capacidad antioxidante y estimula el sistema inmunológico. El selenio es un componente de varias selenoproteínas con función preventiva de algunas formas de cáncer.
En un estudio se enriquecieron hongos ostra con selenio y cinc e introdujeron este alimento en la dieta de animales experimentales para comprobar si la actividad antioxidante y antitumoral se incrementaba. Los resultados mostraron que este hongo enriquecido con selenio y cinc mejoraba la actividad antioxidante y prevenía el desarrollo de cáncer de pulmón en ratones.
Vitaminas
Los hongos se consideran una buena fuente de vitaminas, sobre todo de riboflavina (B2), niacina (B3) y folatos (B9) al ser las que contienen en mayor cantidad. El contenido en riboflavina en los hongos también supera la concentración presente en los vegetales, incluso algunas variedades de hongos presentan concentraciones de riboflavina tan altas como las que se pueden encontrar en el huevo o el queso.
Las vitaminas B1, B12 y C también están presentes en los hongos, aunque en menor cantidad.
Vitamina D: Los hongos son el único alimento no animal que contienen esta vitamina (en forma de precursor) y, por lo tanto, son la fuente natural de vitamina D para los vegetarianos. La vitamina D corresponde a dos compuestos liposolubles: La vitamina D3, colecalciferol, y la vitamina D2 o ergocalciferol. La vitamina D3 es producida en la piel del ser humano y los animales, a partir de u derivado del colesterol ante la exposición a los rayos UVB que recibimos del sol. La vitamina D2 es producida en plantas, hongos y levaduras en forma similar, pero del ergosterol. Ambos compuestos son transformados a nivel hepático (hígado) y renal, siendo estos productos los activos y responsables de las múltiples funciones de la vitamina D en el organismo. Dichas funciones pueden ser agrupadas en:
- Efectos en en el metabolismo de calcio y fósforo. Esto finalmente guarda relación con la integridad de huesos y dientes.
- Efectos en tejidos no calcificados. Se ha establecido que tiene un papel importante en la regulación inmune, función endocrina del páncreas, en la piel, en la función muscular y en el desarrollo cerebral.
Debido a estas funciones se reconocen beneficios del consumo de vitamina D para la prevención o atenuación de algunas enfermedades tales como: artritis reumatoidea, esclerosis múltiple, diabetes tipo 1, enfermedades inflamatorias de la piel, enfermedad inflamatoria intestinal (colitis ulcerosa, enfermedad de crohn), diabetes tipo 2, enfermedades cardiovasculares y en ciertos tipos de cáncer (e.g.: cáncer de colon, de próstata, colorectal, de mama). Debido a estas múltiples funciones, todas relevantes para la salud y al bajo consumo de alimentos ricos en vitamina D (hígado de ciertos peces marinos) y al bajo contenido en la mayoría de los alimentos, se promueve la suplementación y/o la exposición cuidadosa o prudente a la luz solar. De esta manera, el consumir hongos hace un importante aporte a esta vitamina.
EFECTOS EN LA SALUD DEL CONSUMO DE HONGOS COMESTIBLES
Una razón que ha estimulado el consumo de hongos es que se han vuelto atractivos como alimentos funcionales por sus potenciales efectos benéficos sobre la salud humana. Los hongos contienen compuestos bioactivos de alto valor medicinal como lectinas, polisacáridos, compuestos fenólicos y polifenólicos, terpenoides, ergosteroles y compuestos orgánicos volátiles, que son considerados como agentes responsables relevantes para su actividad sobre la salud que incluyen efectos: antitumorales, inmunomoduladoras, antioxidantes, antihipercolesterolemicos, antivirales, antibacterianos, hepatoprotectores y efectos antidiabéticos. Numerosos estudios han revelado que las diferentes especies de hongos son beneficiosas para la prevención y apoyo al tratamiento de varias enfermedades crónicas, como cáncer, enfermedades cardiovasculares, diabetes mellitus y enfermedades neurodegenerativas. Me referiré por ahora preferencialmente a hongos comestibles. Hay otros hongos que tienen un uso más bien medicinal y serán tema para posteos futuros.
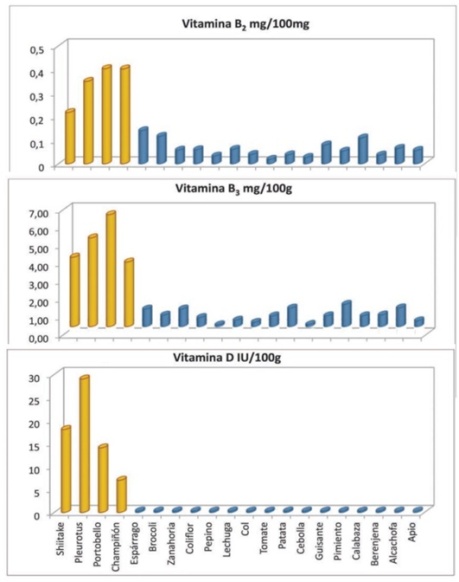
Efecto sobre los niveles de colesterol
Los hongos comestibles son un alimento ideal para la prevención de aterosclerosis debido a su alto contenido de fibra y bajo contenido de grasa. Los hongos, junto a sus extractos, podrían considerarse como una nueva fuente de compuestos con actividad hipocolesterolémica porque son ricos en derivados de ergosterol, eritadenina, beta-glucanos e inhibidores de la enzima HMG-CoA reductasa, la enzima clave en el colesterol endógeno biosíntesis.
La eritadenina, acido (2R,3R)-4-(6-Aminopurin-9-yl)-2,3-dihidroxibutanoico, también conocido como lentisina o lentinacina, es un compuesto con actividad anticolesteronemica. Estudios en ratas muestran que actúa como un inhibidor de la S-adenosilhomocisteína hidrolasa (SAHH), enzima clave en el metabolismo de los fosfolípidos.
La actividad hipocolesterolémica estaría relacionada con otros compuestos presentes en hongos, tales como la fibra dietética y el ergosterol. Este último, se ha identificado como un agente hipocolesterolémico por su capacidad de competir con las moléculas de colesterol por su incorporación en las micelas dietéticas mixtas durante digestión in vitro debido a su similitud estructural.
Compuestos con actividad antioxidante
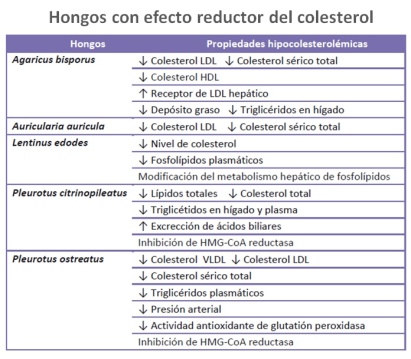
En cualquier organismo metabólicamente activo, los procesos fisiológicos involucran la producción de radicales libres y compuestos reactivos derivados del oxígeno como subproductos. La presencia de estos compuestos más allá de cierto límite, altera el equilibrio antioxidante y provoca estrés oxidativo. Esto conduce a la destrucción de tejidos y células que luego resultan en procesos que afectan el estado de salud, tales como procesos inflamatorios, autoinmunes incluso cánceres. Por lo tanto, mantener el equilibrio oxidativo incorporando antioxidantes en las dietas de los humanos es un factor importante para reducir el daño oxidativo. Compuestos fenólicos, tocoferol, ácido ascórbico y carotenoides son compuestos responsables que se han aislado de las diferentes especies de hongos y se ha informado que además estimulan el sistema inmune, tienen actividad anticancerosa, anti-hipercolesterolémica y actividad antiviral, y mejoran el efecto tóxico de la quimioterapia y radioterapia. Su actividades como antioxidantes ha sido ampliamente estudiada y documentada, especies como Pleurotus, Agaricus, G. lucidum y L. edodes, las que son conocidas por sus profundas actividades antioxidantes. El popular hongo comestible chino Hericium erinaceus se usa para tratar varias dolencias, su extracto de agua caliente exhibe una alta actividad de eliminación de radicales libres que fortalece su uso como nutracéuticos que tienen antioxidantes naturales. Entre los compuestos que tienen actividad antioxidante se encuentran el selenio, la ergotienina, los polisacáridos, compuestos polifenoles y terpenos
Ergotioneina (EGT). Corresponde al compuesto 2-mercaptohistidine trimethylbetaine, aminoácido natural derivado de la histidina con un átomo de azufre en el anillo imidazólico. Es una molécula de tiol/tiona sintetizada solo por algunos hongos y bacterias. Los humanos y algunos animales lo toman de la dieta a través de un transportador especial y se acumula a altos niveles en ciertos tejidos. La ergotioneina no se metaboliza o excreta rápidamente en la orina y está presente en muchos, si no en todos, los tejidos humanos y los fluidos corporales. Tiene potentes propiedades antioxidantes y citoprotectoras in vitro y existe evidencia de que el cuerpo puede concentrarla en sitios de lesión tisular. Se han observado niveles disminuidos de ergotioneina en sangre y/o plasma en algunas enfermedades, lo que sugiere que una deficiencia podría ser relevante para el inicio o la progresión de la enfermedad.
Polisacáridos: compuesto con múltiples efectos
Los polisacáridos son compuestos potentes y comunes derivados de los hongos, ya que exhiben numerosas actividades de beneficios para la salud. Recuerden que los polisacáridos están prsentes como un componente estructural, en la pared celular delos hongos (ver el primer posteo para recordar). Algunos de estos polisacáridos se han desarrollado y explotado como sustancias alimentarias funcionales, incluido el lentinan de Lentinus edodes, el esquizofillan de Schizophyllum commune, el pleuran de la especie Pleurotus, calociban de Calocybe indica y el ganoderan de Ganoderma lucidium. Estos fueron probados y considerados como representantes destacados de beta-glucanos con las cadenas principales de glucosa unidas con enlaces beta (1 → 3) o beta (1 → 6) y distinguidas por diferentes patrones y grados de ramificación. Los monosacáridos más comunes detectados en los polisacáridos de los hongos son glucosa, galactosa, fructosa, xilosa, manosa, fucosa, ramnosa, arabinosa, trehalosa y manitol.
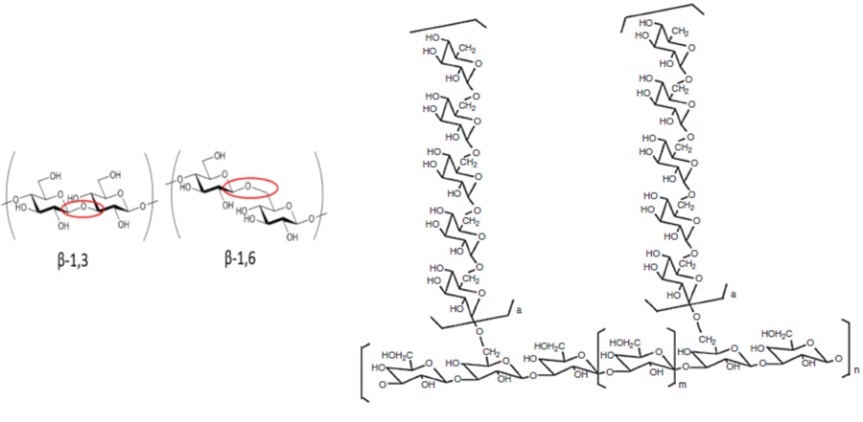
Una gran cantidad de estudios han investigado y revelado las capacidades potenciales de los polisacáridos de hongos en actividades biológicas como muestra la Tabla de más abajo. En general, las actividades antitumorales, inmunomoduladoras y antiinflamatorias de los polisacáridos de hongos son los tres principales beneficios encontrados para la salud. Se estima que la activación de células dendríticas, monocitos, neutrófilos, células asesinas naturales, macrófagos citotóxicos y citocinas (interleucinas, interferones y factores estimulantes de colonias) son los mecanismos de funcionamiento comunes de diferentes polisacáridos de hongos que muestran estas tres actividades.

Por otro lados, los estudios centrados en los beneficios para la salud de los polisacáridos de hongos a través de la regulación de la microbiota intestinal, así como en la función gastrointestinal, han generado un considerable interés por parte de los científicos de todo el mundo. Básicamente, la mayoría de los polisacáridos de los hongos han sido probados como fibras dietéticas y muestran un proceso de interacción con la microbiota colonizada en el tracto gastrointestinal, lo que por lo tanto podría alterar diferentes variaciones de la microbiota intestinal e impactar el nivel de salud del huésped.
RESUMEN
Por lo expuesto, espero que les quede claro que los hongos son un alimento nutricionalmente bueno, especialmente sano. Esto debido a:

Además, por los componentes presentes, se puede decir es un alimento funcional, esto es, tiene beneficios para la salud:

Entonces………. a conseguir buenas recetas…………….. y a comer !!!

PD: no se me ocurre una cancion ad-hoc para este posteo. ¡ se aceptan sugerencias !
NOTA del Redactor : este posteo fue realizado en forma voluntaria, ninguna empresa productora de hongos dio $$$$ para esta actividad.