Sin duda es chocante escuchar que el organismo vivo más grande sea un hongo. Cuando se piensa en los organismos más grandes del planeta, generalmente se visualiza la ballena azul (suelen medir alrededor de 30 m de largo y pueden pesar más de 180 toneladas), o dinosaurios (aunque estos son menores en tamaño que la ballena). Como sea, el récord mundial del organismo viviente más grande lo tiene un hongo. Ahora, no se imagen que hay por ahí una callampa gigantesca o algo así. Se trata de un hongo del genero Armillaria, el cual crece normalmente en el suelo y ataca o destruye árboles.
Se han descritos a la fecha dos trabajos que muestran la gran distribución de superficie (de donde se deduce su gran tamaño) de este tipo de hongos:
- El primero fue publicado en abril de 1992 en la prestigiosas revista Nature por los autores Myron Smith, Johann Bruhn, y Jim Anderson (Smith et al. (1992) identificaron en Michigan un hongo que cubría una superficie de 15 hectáreas, comparable a la de El Pentagono. Bastante grande. ¿no creen? Pueden acceder a la publicacion en cuestion aqui
- El Segundo fue publicado el 2003, por Catherine Parks Brennan Ferguson, Tina Dreisbach Greg Filip, y Craig Schmitt, Wallowa-Whitman, del US Forest Service, identificando en el Parque Nacional de Malheur, Oregon, USA, un hongo que cubría una superficie de 960 hectareas, equivalente a 15 veces el Estadio Nacional…Enorme!! (link)
Datos sobre el género Armillaria.
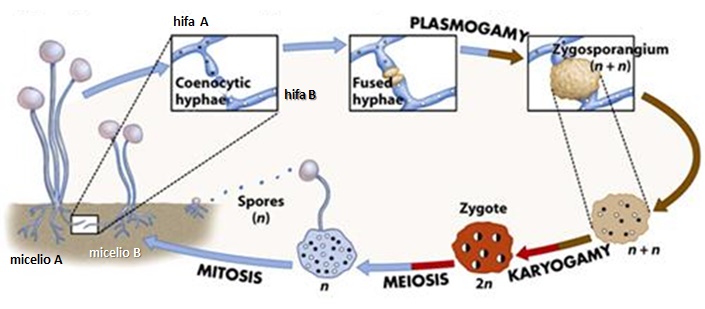
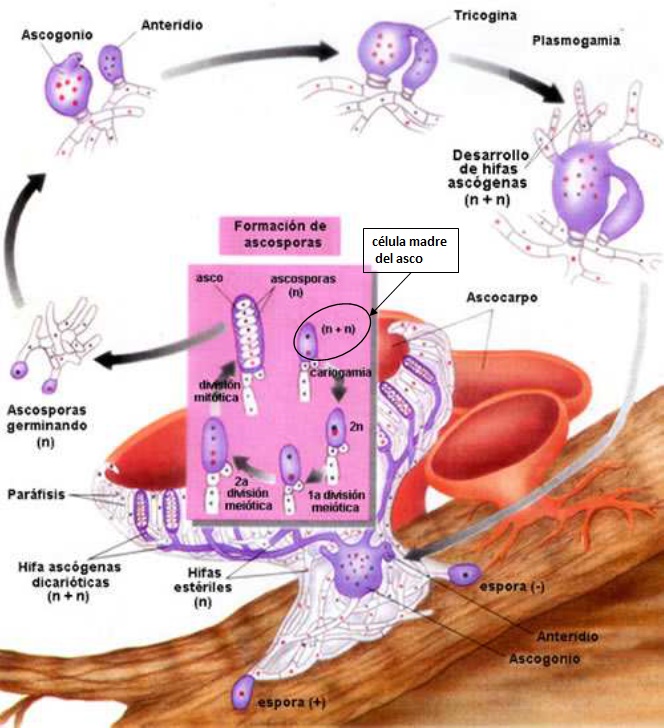
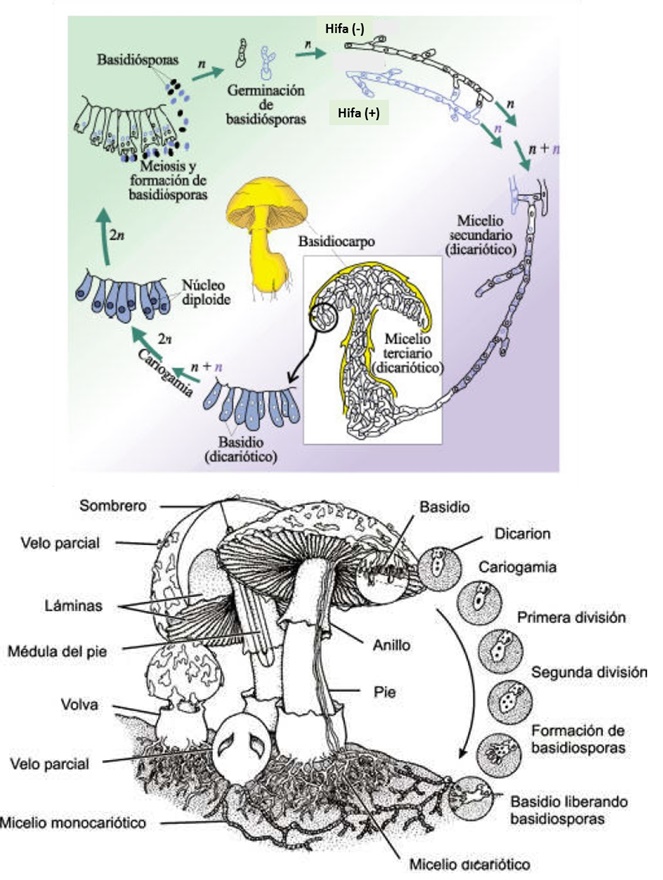
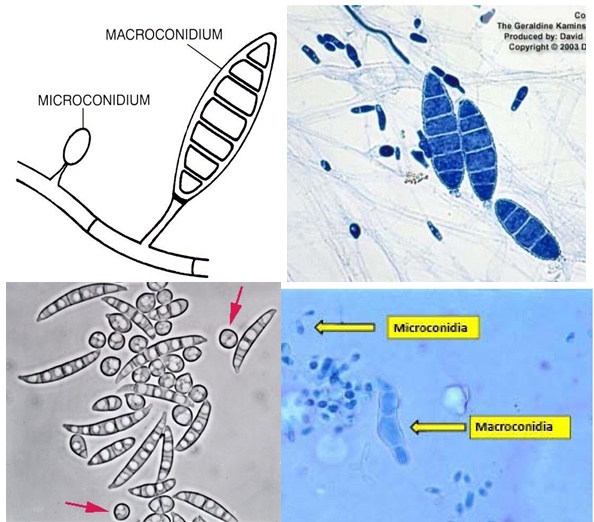
Antes que nada, algunos datos del hongo en cuestión. A los hongos del género Armillaria, se le conoce también como el hongo, seta o champiñón de miel. Corresponden a hongos parásitos basidiomicetes que viven sobre los árboles o arbustos leñosos. Incluye unas 10 especies que antiguamente estaban agrupadas bajo la designación de A. mellea. En las praderas canadienses (particularmente en Manitoba), el término «hongo de la miel» no es muy conocido; ya que debido a la presencia de inmigrantes ucranianos, a menudo el hongo es llamado pidpenky que en ucraniano significa, «debajo del tocón» (el tocón es la parte del tronco de un árbol que queda unida a la raíz cuando lo cortan). Producen masas comestibles de callampas amarillo-café que crecen sobre el suelo. Es su color, más que su sabor, el que le ha dado su nombre de “miel”, a pesar de que algunos dicen que deja un sabor dulce en la boca. Una característica importante del de Armillaria es la formación de rizomorfos, que es una agrupación de hifas en paralelo que se cubre con corteza negra melanizada que le sirve de protección contra el exterior. Los rizomorfos son capaces de transportar nutrientes largas distancias, lo que permite que el hongo crezca a través de áreas pobres en nutrientes ubicadas entre zonas ricas como los tocones. Los rizomorfos actúan como «exploradores» del resto del micelio en la búsqueda de nuevas fuentes de nutrientes. Estos rizomorfos permitirían a las colonias de Armillaria propagarse y llegar a ser bastante grandes.

Armillaria gallica, el primero en ser descrito
El descubrimiento de Smith et al. (1992) se realizó cerca de Crystal Falls, Michigan, próximo a la frontera de Wisconsin. Esta zona había sido en su mayoría bosque nativo (roble, abedul y arce) pero estos se habían reemplazado con pinos (más rentables). Cuando se cortaron los árboles, los tocones se comenzaron a pudrir. Los robles habían sido infectados desde la raíz por Armillaria, pero habían sobrevivido sin dar síntomas ni mortalidad notable. Sin embargo, cuando se plantaron pinos, algunas especies de Armillaria fueron capaces de matar las plántulas jóvenes. La especie particular que obtuvo su atención era Armillaria bulbosa, que ahora se conoce como correcta Armillaria gallica. En dicho el estudio, se colecto micelio vegetativo de Armillaria por medio de una técnica de «cebado», que utiliza varillas de madera de álamo. Dado que Armillaria es un hongo que descompone la madera, el micelio coloniza rápidamente estos cebadores, de donde (una vez colonizada) se puede recolectar fácilmente el micelio del hongo y cultivarlo. También se colectaron esporas directamente de los cuerpos fructíferos y de los rizomorfos. Las muestras se analizaron primero determinado cuan variable eran los tipos de apareamiento (a través de cruzamientos) y mediante técnicas de ADN, observando los patrones de restricción del ADN mitocondrial (ADNm). Tanto los grupos de apareamiento y los patrones de restricción de ADNm son muy variables dentro de las especies de Armillaria. Se incorporaron posteriormente, otros análisis genético-moleculares tales como ensayos de RAPD (ADN polimórfico amplificado al azar) y RFLP (Fragmentos de restricción de largo variable). Estos permiten evaluar la variabilidad individual entre las muestras colectadas. Con esta información, se trazaron mapas de la zona para determinar los límites de cada individuo colectado. Se identificó la presencia de dos clones dentro de todas las muestras. Uno de ellos resulto ser bastante grande, abarcando una superficie de 15 hectáreas. Dentro de esta área, todos los aislados tenían el mismo tipo de apareamiento, el mismo patrón de restricción mDNA, y tenían los mismos once productos de RAPD y cinco marcadores de RFLP. Estos datos indicaron que este clon de 15 hectáreas corresponde a un solo organismo o bien todos ellos eran altamente parecidos. En el trabajo se presentaron argumentos estadísticos que mostraban que la probabilidad de que esto último mencionado era infinitamente pequeño. Teniendo en cuenta que todas las muestras comparten todos los marcadores heterocigotos examinados, la hipótesis más probable es que este clon alcanzó su enorme tamaño a través del crecimiento vegetativo en un determinado tiempo. Mediante la estimación (de manera muy conservadora) de la tasa de crecimiento del hongo en virtud de sus condiciones naturales y extrapolando el peso del clon de muestras de suelo más pequeñas (de nuevo de manera muy conservadora), se estimó que el clon tendría unos 1500 años de edad y pesaba al menos 9.700 kg (casi 100 toneladas), cerca de la masa de un ballena azul adulta.
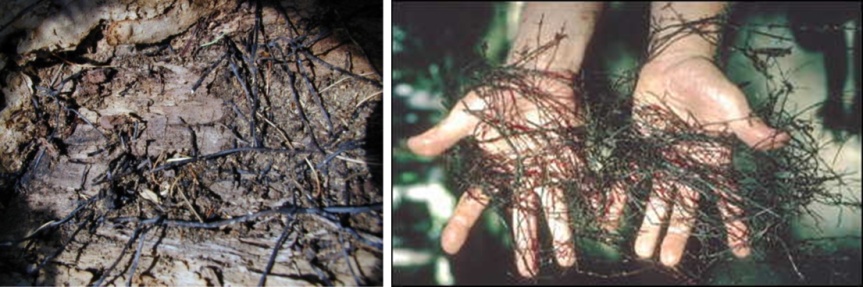
El más grande, Armillaria ostoyae, descrito una década después
Diez años más tarde, Catherine Parks del Servicio Forestal de Estados Unidos en Oregon, junto a otros investigadores, informaron de que habían encontrado un hongo aún mayor, de nuevo Armillaria, pero otra especie, A. ostoyae, en el este de Oregón, dentro del Malheur National Forest. El descubrimiento se hizo luego de que los científicos comenzaran a investigar por qué un gran número de árboles se estaban muriendo. Se colectaron muestras de raíces tomadas de 112 árboles (algunos muertos y otros en decaimiento), los que se analizaron en formar similar a lo descrito en el trabajo de Smith et al (1992). Los resultado mostraron que si bien todos estaban infectados con un solo hongo, A. ostoyae, se podían identificar 5 clones distintos que resultaron ser bastantes grandes. En efectos, estos cubrían superficies que variaron entre 20 y nada menos que 960 hectáreas. A este último clon se le estimó que podría tener al menos unos 2.400 años (aunque incluso podría tener 8.650). Asi, A. ostoyae, no solo es el organismo vivientes más grande, sino que también el más viejo del planeta.

Comentarios
Un aspecto interesante de estos trabajos es el debate científico que se originó acerca de «¿qué es exactamente un organismo?». La mayoría de la gente entiende fácilmente este concepto al pensar en un animal, que tiene límites definidos y es generalmente visible. Sin embargo, gran parte de una planta y más aún en un hongo, el organismo en cuestión no es visible para el ojo desnudo. Para los hongos del suelo, los límites entre individuos son difíciles de establecer. Así, la gran pregunta fue «¿Actúan estos hongos gigantescos como organismos individuales?» Genéticamente al menos, se estableció que los diversos aislados eran idénticos, pero ¿puede una parte del organismo comunicarse con otras partes del organismo? ¿Comparten la fisiología? Si las diferentes partes están creciendo a través de diferentes sustratos ¿se les suministran a las otras partes del hongo los nutrientes que le faltan? Es posible, ya que hay trabajos que demuestran esto (lo comentare más adelante)
Hay que recalcar que a pesar del gran tamaño de los gigantescos micelios de los hongos descritos, los cuerpos fructíferos (callampas, setas) son realmente de tamaño normal (50 a 100 g). Sin embargo, durante la temporada de fructificación, las setas de miel pueden ser muy abundantes en las zonas donde se describieron.
Finalmente, se reconoce que miembros del reino de los hongos son los organismos más grandes en la tierra, entendiendo su enormidad por la gran superficie que han llegado a colonizar y, además, los mas antiguos de la tierra, por el tiempo que se han demorado en establecerse en dicha superficie. Es probable que existan otra Armillaria, aún más grande, por ahí esperando la descubran.
Algo de música relacionada (aunque solo por el nombre del grupo, Proclamation de Gentil Giant)